O’Donnell, Hoare: Bird recovery after pest control
Available on-line at: http://www.newzealandecology.org/nzje/
131
Quantifying the benefits of long-term integrated pest control for forest bird
populations in a New Zealand temperate rainforest
Colin F. J. O’Donnell* and Joanne M. Hoare
Research and Development Group, Department of Conservation, PO Box 11089, Sockburn, Christchurch 8443, New Zealand
*Author for correspondence (Email: [email address])
Published on-line: 16 April 2012
Abstract: The control of introduced mammalian predators has become a standard response to protecting the
viability of threatened wildlife species on oceanic islands. However, examples of successful outcomes of integrated
pest control in forests are few. We investigated the efficacy of a pest control programme in the Landsborough
Valley, New Zealand, during 1998−2009, which used continuous trapping to control mustelids and pulsed aerial
application of the toxin 1080 to control rats (
Rattus spp.) and brushtail possums (
Trichosurus vulpecula). We
predicted recovery in the populations of mohua (
Mohoua ochrocephala) and other predator-sensitive hole-nesting
birds and maintenance of numbers of South Island kaka (
Nestor meridionalis meridionalis). In addition, we
examined whether annual mean counts of mohua and kaka, as potential ‘population indicator species’, could
predict those of other forest bird species. Annual counts of nine species (eight indigenous: bellbird
Anthornis
melanura, brown creeper
Mohoua novaeseelandiae, fantail
Rhipidura fuliginosa, grey warbler
Gerygone
igata, mohua, rifleman
Acanthisitta chloris, tui
Prosthemadera novaeseelandiae and yellow-crowned parakeet
Cyanoramphus auriceps; one introduced: song thrush
Turdus philomelos) showed significant increases during
the 12-year study period. South Island kaka and redpoll (
Carduelis flammea) showed no change with time.
In general, trends in the two focal threatened taxa (mohua and kaka) were poor predictors of trends in other
bird species. Lack of correlation in annual counts between bird species that share a recovery trajectory are
likely due to differences in breeding biology and resource use. Our results suggest that an integrated strategy
for predator management is effective at mitigating the impacts of predation by introduced mammals on forest
birds, including the most vulnerable species, at a landscape scale.
Keywords: 1080; browsers; mast;
Mohoua;
Nestor;
Nothofagus; poisoning; population indicator species;
predators; trapping
Introduction
(Dilks et al. 2003) or pulsed poison operations (Basse et al.
2003), to integrated and sustained control that incorporates a
The introduction of mammalian predators to oceanic islands
number of control techniques (Baber et al. 2009). While there is
has resulted in declines and extinctions of numerous endemic
a growing list of examples of successful pest control operations
species on these islands (Atkinson 1989; Jones et al. 2008).
using trapping or poisoning for single threatened species or
Therefore, the control or eradication of introduced predators
suites of species, the benefits of longer term, integrated pest
has become a major conservation priority in these areas (Towns
control programmes are not well understood.
et al. 2006). Developing effective control techniques can be
An inventory of the forest bird population of the
challenging, particularly when multiple predator species occur
Landsborough Valley was conducted in 1985 as part of a
sympatrically, because of interspecific variability in predator
wider biodiversity assessment of forests in South Westland,
cycles and the need to implement multiple control techniques
New Zealand (O’Donnell & Dilks 1986). The inventory noted
(e.g. Saunders & Norton 2001; Burbidge & Morris 2002;
significant bird populations in the valley, including the largest
Burrows et al. 2003; Donlan & Wilcox 2008).
population of mōhua (yellowhead
Mohoua ochrocephala)
Significant declines in many species of forest birds in
remaining in South Westland and one of the largest populations
response to predation by introduced mammalian predators have
of South Island kākā (
Nestor meridionalis meridionalis) in
been widely documented in New Zealand (reviewed by Innes
the region. The site was recommended for protection, and
et al. 2010). Likewise, the impacts of browsing and predation
subsequently as part of the South Westland World Heritage
by introduced brushtail possums (
Trichosurus vulpecula) on
Area (Department of Conservation 1989). At the time, research
forest structure and threatened species are well documented
indicated that predation by stoats (
Mustela erminea) was a
(O’Donnell 1995; Payton 2000; Sadleir 2000). Based on these
major cause of decline in mōhua populations throughout the
impacts, attempts to control predators and browsers are now
South Island, and that, for populations to be maintained or
widespread in New Zealand forests, even though the benefits
recover, predator control at those sites would be required (Elliott
of control have only been documented for a limited number
1996; O’Donnell 1996a; O’Donnell et al. 1996). Research
of bird and plant species (e.g. Sweetapple et al. 2002; Basse
also indicated that the distribution of South Island kākā was
et al. 2003; Innes et al. 2004; Urlich & Brady 2005; Nugent
shrinking dramatically in the region and that kākā abundance
et al. 2010). Pest control operations in New Zealand comprise
was highest at sites not yet colonised by brushtail possums and
a range of scenarios, from one-off pest specific operations
declined with increasing possum densities (O’Donnell & Dilks
(O’Donnell et al. 1996) and longer term continuous trapping
1986; Rose et al. 1990). Possums were beginning to colonise
New Zealand Journal of Ecology (2012) 36(2): 131-140 © New Zealand Ecological Society.

132
New Zealand Journal of Ecology, Vol. 36, No. 2, 2012
the Landsborough Valley in the early 1980s and control of
Methods
their spread was recommended to protect South Island kākā
and populations of mistletoes (
Peraxilla spp.) from potential
Study area
decline. Mistletoes form a significant seasonal food source
The study was conducted in the mid-Landsborough Valley,
for South Island kākā, but they are also highly palatable to
South Westland, New Zealand (43°S, 169°E; Fig. 1). The
possums and therefore at risk from defoliation (O’Donnell &
study area comprised forested lower valley slopes, fans and
Dilks 1986, 1994; Sweetapple 2008).
terraces on the eastern side of the Landsborough River from
Monitoring of the Landsborough mōhua population
McKerrow Creek in the north to Fraser Creek in the south (c. 7
commenced as part of the Mōhua Recovery Programme
km, 900 ha). The canopy forest is almost entirely silver beech
(O’Donnell 1996b). Numbers dropped from 163 birds in
(
Nothofagus menziesii), with occasional subcanopy Hall’s
1985 to 24 birds in 1991 and 14 birds in 1992 (O’Donnell
totara (
Podocarpus hallii) and rarely kāmahi (
Weinmannia
1996b). This pattern of decline was similar to that observed
racemosa). Large mistletoes (
Peraxilla colensoi,
P. tetrapetala)
in beech forests elsewhere in the South Island (Elliott 1996;
are common epiphytes. The understorey is sparse and the
O’Donnell 1996b; O’Donnell et al. 1996). The significant drop
most common shrubs include broadleaf (
Griselinia littoralis),
in numbers of mōhua detected in 1991 and 1992 followed
Coprosma spp., haumakaroa (
Raukaua simplex), fuchsia,
heavy beech masting and subsequent irruptions in rodent and
fivefinger (
Pseudopanax arboreus) and weeping mapou
stoat populations across South Island beech forests (O’Donnell
(
Myrsine divaricata). Ground cover is dominated by leaf litter
1996b; O’Donnell & Phillipson 1996).
and beds of moss.
Concerns for maintaining plant communities in the
Landsborough Valley led to a control programme for possums
Plant phenology
being initiated by the Department of Conservation (DOC) in
1994. As concerns for the continued viability of the mōhua and
The amount of beech seedfall in a year is a clear indicator of
kākā populations increased, the possum-control programme
predator levels and predation risk over the subsequent year
was integrated with continuous trapping of stoats and rodents
(King 1983; O’Donnell & Phillipson 1996) and beech seeding
in 2000. The integrated programme aimed to control sufficient
indicates that food resources are sufficient for kākā to breed
numbers of stoats, rodents and possums to maintain populations
(O’Donnell & Dilks 1994; Moorhouse 1997). Therefore,
of forest birds and possum-palatable species (e.g. mistletoe,
because the flowering intensity of silver beech (and the
fuchsia
Fuchsia excorticata,
Raukaua spp. and
Pseudopanax
resultant seedfall) varies considerably from year to year, the
spp.).
phenology of silver beech was monitored over the duration of
The primary objective of this study was to monitor the
long-term response of forest birds to integrated pest control (a
combination of ground trapping for stoats and aerial poisoning
aimed at controlling brushtail possums and rats (
Rattus spp.)).
We predicted that if integrated pest control was effective we
should detect: (1) recovery in numbers of mōhua and other
hole-nesting birds; (2) maintenance of the high South Island
kākā numbers in the study area (as opposed to dramatic
declines documented in other parts of South Westland); and
(3) a general recovery of forest bird numbers.
Because threatened species receive a disproportionate
share of conservation spending and attention compared with
common species, it is useful to know whether (and how) their
management contributes indirectly to the preservation of other
species (Bonn et al. 2002; Tognelli 2005). The presence of
mōhua (Nationally Vulnerable; Miskelly et al. 2008) and South
Island kākā (Nationally Endangered; Miskelly et al. 2008) in
our study area provided an opportunity to examine their use
as population indicator species (a species whose population
trends can be used as an index of trends in other species; Hoare
et al. 2010) for a range of other forest birds in a situation with
a shared ecological driver (control of introduced mammalian
predators). Threatened species, by definition, are the species
most vulnerable to threat processes and thus can function well
as indicators of trends in biodiversity generally (Lawler et al.
2003; Warman et al. 2004; Tognelli 2005; Larsen et al. 2007).
However, despite many studies that target threatened and rare
species to set priorities for land conservation, we found no
evidence of threatened species being evaluated as population
indicator species in New Zealand. As a result we also examined
the degree to which annual counts of mōhua and South Island
kāka could be used to predict counts of other forest birds.
Figure 1. Location of the Landsborough Valley, South Island,
New Zealand.
O’Donnell, Hoare: Bird recovery after pest control
133
the study at Patsy Creek in the lower Landsborough Valley.
estimate of absolute abundance, and as such these counts
Beech seedfall typically occurs in autumn (February–March)
are incomplete counts, for which we could not correct for
through to early spring, with the majority of seed falling in
the probability of detection. However, indices of relative
March−May (Wardle 1984). Seedfall was monitored from
abundance
such as this are useful when investigating large-
March to May each year using the standardised New Zealand
scale trends in multi-species assemblages over moderate
protocols (
sensu Wardle 1984). Seeds were collected in standard
time frames (Thompson et al. 1998; Hutto & Young 2003;
funnels (plastic funnels 0.28 m in diameter, Gyro Plastics,
Johnson 2008). There are also practical advantages to using
Lower Hutt) placed along a random line-transect (8 funnels,
this technique in New Zealand rainforests (and elsewhere)
1.25 m off the ground, 50 m apart, > 50 m away from a forest
where considerable limitations have been encountered when
edge or 20 m away from a canopy gap). Seeds that fell into
applying techniques that require the calculation of detection
funnels were collected in stockings fitted over the narrow ends
probabilities to multiple species with different behavioural
of the funnels. Funnels were opened on 1 March and closed
responses (Hutto & Young 2003; Greene et al. 2010; T. Greene,
on 31 May each year. At the end of the collecting period, all
DOC, Christchurch, pers. comm.).
seeds were counted and the number of seeds per square metre
Counts were undertaken over three consecutive days by
was calculated. Wardle (1984) defined masting events as: full
a team of three expert bird counters. All bird species seen
(or heavy) = >4000 seeds m–2, partial = 500–4000 seeds m–2,
and heard within 5-min periods were counted at 112 count
poor = <500 seeds m–2.
stations. Counts were carried out in standardised fine conditions
between 0900 and 1500 hours to avoid times of the day when
Pest control
conspicuousness of birds is most variable (dawn and dusk). To
Three techniques for controlling mammalian pests have been
ensure independence, stations were spaced at 200-m intervals
used at the study site: standard Fenn trapping (King et al. 1994)
along a transect through habitats representative of the study
aimed at stoats; ground control (trapping and poisoning) aimed
area (Dawson 1975). The transect followed the forest edge
at brushtail possums; and aerial bait application using baits
from McKerrow Creek down the valley to Fraser Creek, then
impregnated with the toxin sodium fluoroacetate (compound
turned inland and ran back up the valley, parallel with the
1080) aimed primarily at possums and rats.
edge but 200−500 m into the forest. One count per station
The first operation (8585 ha) to control possums in the
was conducted, except for the central 65 stations, which were
study area involved ground trapping (using baited Victor traps
counted twice on each occasion (
n = 177 counts).
set along bush edges) and poison baiting (using cyanide baits
on ridges), and was carried out in January–March 1994. Since
Data analysis
then, possum control has been by periodic aerial application
We used generalised linear mixed-effects models to investigate
of 1080-laced baits (8-g cereal pellets, 0.15% w/w 1080 RS5
whether trends in bird counts were explained by time (year)
bait) sown at 2 kg ha–1 after a prefeed drop of non-toxic 6-g
and amount of beech seedfall (seeds m–2 year–1) for each bird
cereal pellets (RS5) at 1 kg ha–1. Toxin drops centred on the
species counted. Models contained counts as the response
study area were undertaken in July 1998 (7340 ha), June 2000
variable, year and seedfall as predictor variables, and count
(3750 ha), December 2004 (16 420 ha) and October 2009
station as a random effect to account for the repeated measures
(12 170 ha). The timing of these toxin drops was triggered
nature of the study. Individual models for each species were
by increases in trap-catch indices for possums (and, in 2009,
fitted using a Poisson distribution. We used the statistical
rats) and the occurrence of full beech mast in the autumn
programme R (version 2.12.1; R Development Core Team
before control (T. Farrell, DOC, Hokitika, R. Suggate, DOC,
2010) for all analyses. Predator captures from Fenn-trapping
Christchurch, pers. comm.).
were not included in the model because seedfall is the best
Stoats were targeted using Fenn trapping continuously
predictor of likely predation pressure during the following bird
throughout the year from 2000, with trap checks and trap
breeding season (King 1983; O’Donnell & Phillipson 1996)
clearing usually occurring at 3-monthly intervals. Two Fenn
and an unknown number of predators (likely the majority)
traps were installed in each of 93 tunnels placed at 200-m
would have been killed following toxin applications.
intervals along transects throughout the study area (along the
We then used Bayesian techniques to evaluate the two
bird counting line described below) following best practice
threatened species (mōhua and South Island kākā) as potential
(
sensu King et al. 1994). Tunnels were baited with hen’s eggs.
indicators of population trends for every other species, by
fitting a multivariate linear regression model with an AR1
Bird counts
autoregressive structure. For these analyses we used only the
In 1998, bird monitoring by counts along 31 line-transects
continuous data series from 2002 to 2009 (i.e. counts from
ceased and was changed to a more intensive regime of point
1998 were excluded). Bayesian inference via Markov chain
counts to increase the power of the monitoring programme
Monte Carlo (MCMC) methods allows greater flexibility in
to detect changes in abundance. Standardised five-minute
model fitting compared with maximum-likelihood analyses
point counts (
sensu Dawson & Bull 1975; Dawson 1981)
(Link et al. 2002; Gelman et al. 2004; Royle & Dorazio
were undertaken in November in 1998 and annually from
2006), including the ability to model temporal dependence.
November 2002 to 2009.
Specifically, we assumed that the observations were drawn
Worldwide, the point-count method is the most widely
from a multivariate normal distribution with mean of the
ith
used technique for counting birds (Thompson et al. 1998),
species and
jth time period given by
μij and variance-covariance
and is commonly used for counting songbirds in Europe, the
matrix given by
Τ. The
μij were related to the indicator
USA and New Zealand (e.g. Clout & Gaze 1984; Miller &
species counts
Xj according to the linear model
μij =
β0
i +
βl
iXj.
Anderson 1992; Bibby et al. 2000; Innes et al. 2004). Point
We assumed non-informative normal priors for the beta terms
counts provide information on the relative abundance
of a
(
β0i ~
N(0, 106) and
β1i ~
N(0, 106)) for all indicator species. The
population (indices of relative abundance), rather than an
variance-covariance matrix (
T) was structured to reflect an AR1

134
New Zealand Journal of Ecology, Vol. 36, No. 2, 2012
autoregressive power correlation structure. The covariance of
Results
successive observations over time was described by cov(
yij,
yij+1) =
ρ/ τ where
ρ is the correlation coefficient and
τ is the
Seedfall and predator trap catch
inverse of the sample variance. We assumed a uniform prior
Silver beech seedfall was recorded in all years, with partial
on
ρ from 0 to 1, and a uniform distribution for the prior on
mast or full mast occurring in five of the 11 years (Fig. 2).
τ from 0 to 100.
There were 118 rounds of trap checking in the study area
We fitted the models in the software WinBUGS (version
during the 10 years (2000–2010) of the study period, resulting
1.4.3; Imperial College and MRC, UK). After convergence,
in the trapping of 806 predators representing three species.
a further 10 000 iterations were used for inference. Positive
The majority were stoats (
n = 608, 75.4%), with most of the
and negative correlations between species were identified
remainder being ship rats (
Rattus rattus;
n = 189, 23.5%) and
using 95% credible intervals on the slope (
β1
i) terms for which
a few mice (
Mus musculus;
n = 9, 1.1%). There was marked
the limits excluded zero. Positive trends are defined as those
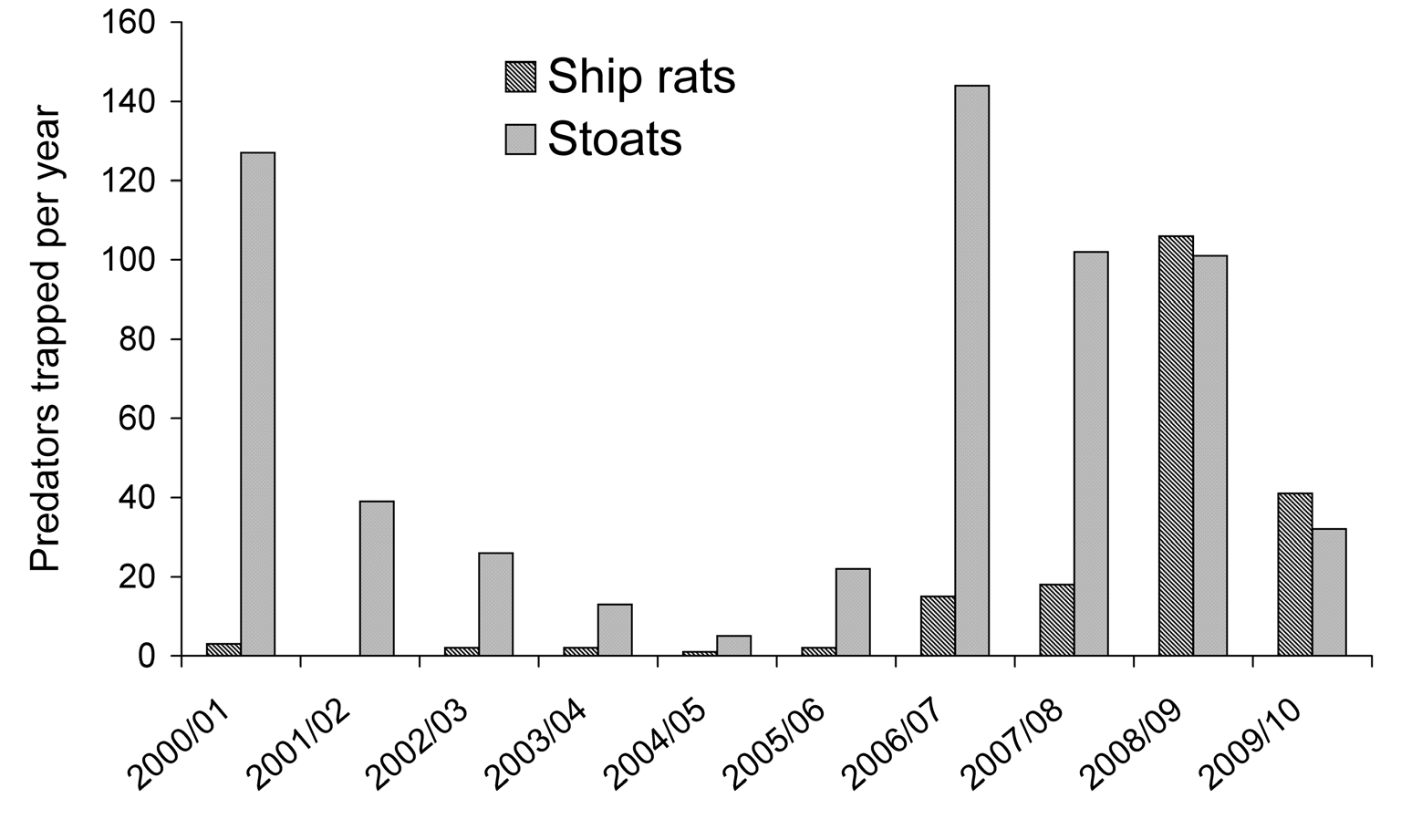
variation in annual captures of predators. Stoats were caught
in which interannual changes are in the same direction and
in relatively high numbers in four of the 10 years (Fig. 3).
negative trends are those that are divergent.
Three of these years coincided with partial or full beech mast
(2000, 2006, 2008; Fig. 2), but captures were low after the
partial mast of 2003.
–2
Seeds/m
Figure 2. Density of beech seedfall (seed m–2), Patsy Creek, Landsborough Valley, March–May inclusive, 1999−2009.
Figure 3. Annual trap catches of stoats (
Mustela erminea) and ship rats (
Rattus rattus) in the Kea Flat study area, Landsborough
Valley, November–October inclusive, 2000−2010 (
n = 93 tunnels each with 32 traps, uncorrected for sprung traps).
O’Donnell, Hoare: Bird recovery after pest control
135
Trends in bird counts
Counts for seven bird species (blackbird, mōhua, redpoll,
Between 168 and 177 bird counts were undertaken annually,
rifleman, silvereye, tomtit and yellow-crowned kakariki) were
except in 2006 when counting was curtailed by poor weather
higher in years when seedfall was also high (Table 1). Fewer
(
n = 113 counts). We analysed counts for 15 bird species (see
bellbird and South Island kākā were recorded in years with
Table 1 for scientific names) that were counted >100 times
high seedfall (Table 1).
during all counts at all stations (Fig. 4).
Eight indigenous
Trends in the mean annual counts of South Island kākā
species (bellbird, brown creeper, fantail, grey warbler, mōhua,
in the Landsborough Valley were negatively correlated with
rifleman, tūī and yellow-crowned parakeet) and one introduced
trends in mōhua, tomtits and yellow-crowned parakeets (Fig.
species (song thrush) showed significant (
P < 0.05) increases
5). Trends in the mean annual counts of mōhua were positively
between 1998 and 2009 (Fig. 4, Table 1). South Island kākā and
correlated with riflemen and yellow-crowned parakeets (Fig.
redpoll showed no change with time. Two indigenous species
6). Mean annual counts in other taxa were not correlated with
(silvereye, tomtit) and two introduced species (blackbird,
either South Island kākā or mōhua counts (Figs 5 & 6).
chaffinch) showed a significant decline (Table 1).
1998 2000 2002 2004 2006 2008
1998 2000 2002 2004 2006 2008
Bellbird
Blackbird
Brown creeper
Chaf inch
Fantail
1.5
1.0
0.5
0.0
Grey warbler
Kaka (South Island)
Mohua
Redpoll
Rifleman
1.5
)
ount
1.0
ean C
M
0.5
log (
0.0
Silvereye
Song thrush
Tomtit
Tui
Yellow-crowned parakeet
1.5
1.0
0.5
0.0
1998 2000 2002 2004 2006 2008
1998 2000 2002 2004 2006 2008
1998 2000 2002 2004 2006 2008
Fig. 4
Year
Figure 4. Annual mean counts of birds per five-minute count in the mid-Landsborough Valley, 1998−2009. Trends were significant
(
P < 0.05) for all bird species except South Island kākā and redpoll. See Table 1 for scientific names of birds.
33
Table 1. Influence of year and seedfall on trends in bird counts in the Landsborough Valley, 1998–2009. Generalised linear
mixed-effects models contained counts as the response variable, year and seedfall as predictor variables, and count station
as a random effect (* = significant at
P < 0.05).
__________________________________________________________________________________________________________________________________________________________________
Species
Scientific name
Year
Seeds
Z-value
P-value
Z-value
P-value
__________________________________________________________________________________________________________________________________________________________________
Bellbird
Anthornis melanura
10.924
<0.001*
−1.241
0.022*
Blackbird
Turdus merula
−8.378
<0.001*
5.124
<0.001*
Brown creeper
Mohoua novaeseelandiae
9.307
<0.001*
0.729
0.466
Chaffinch
Fringilla coelebs
−7.805
<0.001*
1.205
0.228
Fantail
Rhipidura fuliginosa
2.271
0.023*
0.388
0.698
Grey warbler
Gerygone igata
2.289
0.022*
0.548
0.584
Kākā (South Island)
Nestor m. meridionalis
0.539
0.590
−7.126
<0.001*
Mōhua
Mohoua ochrocephala
15.331
<0.001*
4.875
<0.001*
Redpoll
Carduelis flammea
−0.833
0.404
3.333
<0.001*
Rifleman
Acanthisitta chloris
6.018
<0.001*
2.044
0.041*
Silvereye
Zosterops lateralis
−5.120
<0.001*
2.898
0.003*
Song thrush
Turdus philomelos
3.987
<0.001*
−0.244
0.807
Tomtit
Petroica macrocephala
−5.936
<0.001*
6.426
< 0.001*
Tūī
Prosthemadera novaeseelandiae 16.590
<0.001*
−1.290
0.179
Yellow-crowned parakeet
Cyanoramphus auriceps
4.869
<0.001*
18.644
< 0.001*
__________________________________________________________________________________________________________________________________________________________________
F 136
ig. 5
New Zealand Journal of Ecology, Vol. 36, No. 2, 2012
Figure 5. Correlations in population trends (as measured
Bellbird
by fitted slopes from an AR1 model) based on raw annual
count data between South Island kākā and other forest bird
Blackbird
populations in the mid-Landsborough Valley. Population
Brown creeper
correlations (those for which the 95% credible intervals do
not overlap zero) are indicated with an asterisk. See Table
Chaf inch
1 for scientific names of birds.
Fantail
Grey warbler
Mohua
*
Redpoll
Rifleman
Silvereye
Song thrush
Tomtit
*
Tui
Yellow-crowned parakeet
*
Fig. 6
-6
-4
-2
0
2
Value
Figure 6. Correlations in population trends (as measured
Bellbird
by fitted slopes from an AR1 model) based on raw annual
Blackbird
count data between mōhua and other forest bird populations
in the mid-Landsborough Valley. Population correlations
Brown creeper
(those for which the 95% credible intervals do not overlap
zero) are indicated with an asterisk. See Table 1 for scientific
Chaf inch
names of birds.
Fantail
Grey warbler
Kaka (South Island)
Redpoll
Rifleman
*
Silvereye
Song thrush
Tomtit
34
Tui
Yellow-crowned parakeet
*
-0.5
0.0
0.5
1.0
1.5
Value
Discussion
forests as a result of predation by introduced rats and stoats
(Gaze 1985; O’Donnell & Dilks 1986; Elliott 1996; Dilks
Response of birds to management
et al. 2003). Declines have been reversed at a local scale (<
Relative abundance of the majority of forest birds in the
100 ha) through intensive trapping of stoats (O’Donnell et al.
Landsborough Valley changed following the implementation
1996; Dilks 1999). However, when these techniques were
of integrated pest control, with increases in nine species and
applied at a landscape scale (13 000 ha), but did not include
no decreases in abundance in another two species during the
specific rat control, declines continued (Dilks et al. 2003). In
period of study. The increase in the relative abundance of
contrast, the translocation of mōhua to predator-free offshore
mōhua, one of the two species most vulnerable to predator
islands has been highly successful, with large populations now
impacts, was particularly notable. Dramatic declines in mōhua
established on eight islands (R. Cole, DOC, Invercargill, pers.
have occurred over the last 40 years in most South Island
comm.). Mōhua counts in the Landsborough Valley increased
35
O’Donnell, Hoare: Bird recovery after pest control
137
from a low of 14 birds encountered when the count stations
species in their response to the integrated pest management
were established in 1992 (O’Donnell 1996b) to a cumulative
strategy. Based on the increased abundance of mōhua and the
sum of >300 birds recorded on the five-minute counts during
absence of any decline in South Island kākā in the Landsborough
2007−2009.
(contrasting with ongoing declines elsewhere), the pest control
In the Landsborough Valley, the forecast decline in numbers
strategy deployed appeared to be sufficient to mitigate the
of South Island kākā (Rose et al. 1990; Veltman 2000) did
impacts of predation on populations of both species. However,
not eventuate, and we suggest this may in part be due to the
we are unable to apportion increased abundance of mōhua to
protection from predation provided by the pest control operation
a particular component of the pest control programme, largely
in the valley. We did not necessarily expect South Island
because stoats and rats are either killed directly by trapping
kākā numbers to increase dramatically in the Landsborough
or indirectly through secondary poisoning, and rats are also
Valley over the study period, firstly, because a proportion
killed by directly-targeted poison operations (Murphy et al.
of the population was likely to spend some time outside of
1998, 1999).
the valley and, secondly, because they have a relatively low
The predictive framework using beech seedfall as an
reproductive rate making them slow to respond to benefits of
indicator for determining when different forms of pest
pest control. Kākā in South Westland are sequential specialist
control were required was useful in the Landsborough Valley.
foragers, moving extensively across the landscape, exploiting
Generally, though not exclusively, there was a strong positive
seasonal food sources (O’Donnell & Dilks 1989, 1994). Thus,
relationship between heavy beech seedfall and the occurrence
although South Island kākā are particularly prone to predation
of peaks in predator captures, as has been found in other studies
on the nest by stoats, their abundance and survival reflects the
(King 1983; O’Donnell & Phillipson 1996). Heavy seedfall
distribution of threats in the wider landscape.
also coincided with intensive flowering of
Peraxilla mistletoes
We cannot account for declines in four bird species.
in the summers of 1999/2000, 2005/06 and 2008/09 and there
Blackbirds, chaffinches and tomtits tend to feed on or close
was moderate flowering in 2002/03 (P. Knightbridge, DOC,
to the ground (O’Donnell & Dilks 1994) so may be more
Hokitika, pers. comm.). Pulses in trap-catch rates of stoats
vulnerable to predation compared with the other species
associated with full beech mast (> 4000 seeds m–2; Wardle 1984)
monitored (e.g. Brown et al. 1998). It is also possible that
are usually associated with real irruptions in populations, but
factors such as competition influence the persistence of species
there is uncertainty as to whether partial masts have the same
when overall bird numbers increase (Innes et al. 2010). In this
effect (O’Donnell & Phillipson 1996). In the Landsborough
case, the dramatic increases in the two aggressive honeyeater
Valley, the partial mast in 2006 did not result in increased stoat
species (bellbird and tūī) may influence persistence of the
captures; a similar outcome to that observed in the Eglinton
third species that consumes similar nectar foods (silvereye)
Valley (O’Donnell & Phillipson 1996). However, the partial
(O’Donnell & Dilks 1994).
mast in 2008 did. There was a high capture rate for stoats
Many bird species appeared to benefit from beech seed
in 2007 when there was virtually no seedfall, but there was
production, as evidenced by higher bird counts in years with
no increase in captures following the mast seeding in 2009.
high seedfall. These responses are likely to relate to increases in
Based on the low capture rate in 2009, we infer that the 1080
productivity or survival, which reflect increased availability of
control operation in 2009 fulfilled its objective; this was the
invertebrate foods (e.g. Alley et al. 2001). The effect appeared
first season in which the application of aerial 1080 control in
greatest in yellow-crowned parakeet, which breeds prolifically
the study area was timed specifically to maximise control of
following beech masting in response to the abundance of seed
predators, particularly rats.
(Elliott et al. 1996).
Two risks to non-target indigenous species have been
identified when using toxins to control introduced pests: killing
Efficacy of the integrated pest management strategy
non-target species directly if they consume baits, or indirectly
We could not practically replicate monitoring in a comparable
through secondary poisoning (Spurr 1979; Notman 1989;
non-treatment area in this study, nor was pre-treatment
Lloyd & McQueen 2002; Powlesland et al. 2003; Westbrooke
monitoring undertaken using five-minute bird counts. In future
& Powlesland 2005). The majority of bird species in the
monitoring studies it would be prudent to adopt a ‘Before-after
Landsborough Valley were unlikely to encounter either toxic
Control-impact (BACI)’ experimental design to strengthen the
baits or invertebrates that may have fed on baits because they
potential inference of studies such as this one. Nonetheless,
primarily forage in the upper strata of the forest, particularly
we suggest that the overall increases in bird abundance were
the canopy (O’Donnell & Dilks 1994). We conclude that on the
likely to be a result of successful implementation of integrated
four occasions where 1080 was used in the study area, there
pest control. Declines in the Landsborough Valley before
were no significant negative impacts on indigenous birds at the
control started, as well as declines in some forest bird species
population level, largely because indices for most indigenous
reported in similar forest types elsewhere in New Zealand
species increased following the operations.
where pest management has not been implemented (e.g. Elliott
1996; McLennan et al. 1996; Dilks et al. 2003; Elliott et al.
Population indicator species
2010), contrast with results from this study. For example,
Annual mōhua counts were positively correlated with two
Elliott et al. (2010) analysed a 30-year point-count monitoring
species, rifleman and yellow-crowned parakeet, despite
dataset of forest birds in an unmanaged beech forest at Nelson
sharing positive linear trends with a further six species
Lakes, New Zealand, and found a significant change in the
over the duration of the study. We suggest that correlations
bird community structure, with five common native species
between mōhua, rifleman and yellow-crowned parakeet may
(bellbird, rifleman, grey warbler, tomtit, tūī) declining in
have resulted from peak breeding in both species occurring
abundance. Declines were attributed to ongoing impacts of
simultaneously following heavy seedfall (Elliott et al. 1996).
predation.
Furthermore, breeding success is likely to have been enhanced
Mōhua and South Island kākā, as the two threatened taxa in
by controlling predator irruptions that usually coincide with
the Landsborough Valley, were likely to be the most sensitive
peak breeding. Mōhua and yellow-crowned parakeets are
138
New Zealand Journal of Ecology, Vol. 36, No. 2, 2012
known to suffer significant predation on incubating females,
dwelling arthropods and house mice to beech seeding
eggs and chicks by introduced mammals during the summers
in the Orongorongo Valley, New Zealand. Journal of the
following heavy seedfall where predators are not controlled
Royal Society of New Zealand 31: 425–452.
(Elliott et al. 1996; O’Donnell et al. 1996; Dilks et al. 2003)
Atkinson IAE 1989. Introduced animals and extinctions. In:
and this is also likely to be true for riflemen, based on recent
Western D, Pearl MC ed. Conservation for the twenty-first
declines (Miskelly et al. 2008; Elliott et al. 2010).
century. New York, Oxford University Press. Pp. 54–75.
Counts of South Island kākā were negatively correlated
Baber M, Brejaart R, Babbitt K, Lovegrove T, Ussher G 2009.
with counts of mōhua, tomtit and yellow-crowned parakeet,
Response of non-target native birds to mammalian pest
which may be attributable either to their asynchrony in how
control for kokako (
Callaeas cinerea) in the Hunua Ranges,
each species responds to food availability and use or to their
New Zealand. Notornis 56: 176–182.
relatively slow reproductive rates. Increases in the productivity
Basse B, Flux I, Innes J 2003. Recovery and maintenance
of kākā associated with increased food supplies occur in the
of North Island kokako (
Callaeas cinerea wilsoni)
season prior to heavy seedfall (Moorhouse et al. 2003), whereas
populations through pulsed pest control. Biological
productivity of yellow-crowned parakeet increases significantly
Conservation 109: 259–270.
in the breeding season following masting (Elliott et al. 1996).
Bibby CJ, Burgess ND, Hill DA, Mustoe SH 2000. Bird census
Positive relationships between high seedfall and counts of both
techniques. 2nd edn. London, Academic Press. 302 p.
mōhua and tomtits (Table 1) suggest that productivity in these
Bonn A, Rodrigues ASL, Gaston KJ 2002. Threatened and
species also follows beech masting.
endemic species: are they good indicators of patterns
The inability of the two threatened taxa (mōhua and
of biodiversity on a national scale? Ecology Letters 5:
South Island kākā) to predict trends in the more common bird
733–741.
species is likely to be a result of their particular behavioural
Brown KP, Moller H, Innes J, Jansen P 1998. Identifying
and breeding strategies. Compared with other forest birds,
predators at nests of small birds in a New Zealand forest.
mōhua are among the most susceptible species to predation,
Ibis 140: 274–279.
because their peak in breeding overlaps precisely with peak
Burbidge AA, Morris KD 2002. Introduced mammal
abundance of stoats and rats, and female mōhua demonstrate
eradications for nature conservation on Western Australian
no predator avoidance behaviours when incubating (Elliott
islands: a review. In: Veitch CR, Clout MN eds Turning
1996; O’Donnell et al. 1996). Kākā, however, appear to
the tide: the eradication of invasive species. Gland,
anticipate masting events and breed prior to the peak in seedfall
Switzerland, IUCN. Pp. 64–70.
(Moorhouse et al. 2003). Thus, the extent to which annual
Burrows ND, Algar D, Robinson AD, Sinagra J, Ward B,
changes in abundance of these two threatened taxa should be
Liddelow G 2003. Controlling introduced predators in
expected to vary in synchrony with a range of other taxa is
the Gibson Desert of Western Australia. Journal of Arid
debatable. Common species, selected on the basis of shared
Environments 55: 691–713.
traits, may be better indicators of a more widespread response
Clout MN, Gaze PD 1984. Effects of plantation forestry on
to management. Annual variations in abundance of birds are
birds in New Zealand. Journal of Applied Ecology 21:
likely to be more extreme in beech forests because heavy
795–815.
seedfall results in extreme variability in both food supplies
Dawson DG 1981. Counting birds for a relative measure
and predator densities. Our results demonstrate that multiple
(index) of density. In: Ralph CJ, Scott JM eds Estimating
species can benefit from integrated pest control, but that the
numbers of terrestrial birds. Studies in Avian Biology 6.
pattern of response varies among species. This raises the issue
Cooper Ornithological Society. Pp. 12–16.
of the scale at which population indicator relationships should
Dawson DG, Bull PC 1975. Counting birds in New Zealand
be evaluated and applied to outcome monitoring programmes.
forests. Notornis 22: 101–109.
Department of Conservation 1989. Nomination of South-West
New Zealand (Te Wāhipounamu) by the Government of
Acknowledgements
New Zealand for inclusion in the World Heritage List.
Wellington, Department of Conservation.
Thank you to Megan Hieatt, John Lyall, Karen Mayhew, Glen
Dilks P 1999. Recovery of a mohua (
Mohoua ochrocephala)
Newton, Josh Stephens, Paul van Klink and Ron van Mierlo
population following predator control in the Eglinton
for conducting bird counts; Adrian Monks and Ian Westbrooke
Valley, Fiordland, New Zealand. Notornis 46: 323–332.
for statistical advice; Rebecca Wilson, Mark Martini and
Dilks P, Willans M, Pryde M, Fraser I 2003. Large scale stoat
Phil Knightbridge for providing data on pest control and
control to protect mohua (
Mohoua ochrocephala) and kaka
plant phenology; and Lynette Clelland, Graeme Elliott, Phil
(
Nestor meridionalis) in the Eglinton Valley, Fiordland,
Knightbridge, John Lyall, Helen Otley, Tim Shaw, Ron van
New Zealand. New Zealand Journal of Ecology 27: 1–9.
Mierlo and the journal referees for useful discussions and
Donlan CJ, Wilcox C 2008. Diversity, invasive species and
constructive comments on drafts of this manuscript. Funding
extinctions in insular ecosystems. Journal of Applied
for the analysis of these data was provided by the Cross
Ecology 45: 1114–1123.
Departmental Research Pool (Department of Conservation
Elliott GP 1996. Mohua and stoats: a population viability
Research Investigation 3940).
analysis. New Zealand Journal of Zoology 23: 239–247.
Elliott GP, Dilks PJ, O’Donnell CFJ 1996. The ecology of
yellow-crowned parakeets (
Cyanoramphus auriceps)
in
Nothofagus forest in Fiordland, New Zealand.
References
New Zealand Journal of Zoology 23: 249–265.
Elliott GP, Wilson PR, Taylor RH, Beggs JR 2010. Declines in
Alley JC, Berben PH, Dugdale JS, Fitzgerald BM, Knightbridge
common, widespread native birds in a mature temperate
PI, Meads MJ, Webster RA 2001. Responses of litter-
forest. Biological Conservation 143: 2119–2126.
O’Donnell, Hoare: Bird recovery after pest control
139
Gaze PD 1985. Distribution of yellowheads (
Mohoua
Taylor GA 2008. Conservation status of New Zealand
ochrocephala) in New Zealand. Notornis 32: 261–269.
birds, 2008. Notornis 55: 117–135.
Gelman A, Carlin JB, Stern HS, Rubin DB 2004. Bayesian
Moorhouse R, Greene T, Dilks P, Powlesland R, Moran L,
data analysis. 2nd edn. Boca Raton, FL, Chapman &
Taylor G, Jones A, Knegtmans J, Wills D, Pryde M,
Hall/CRC. 668 p.
Fraser I, August A, August C 2003. Control of introduced
Greene T, Jones A, Dennis G, Sachtleben T 2010. Distance
mammalian predators improves kaka
Nestor meridionalis
sampling to determine kaka (
Nestor meridionalis
breeding success: reversing the decline of a threatened
septentrionalis) density within Waipapa Ecological Area,
New Zealand parrot. Biological Conservation 110: 33–44.
Pureora. New Zealand Journal of Ecology 34: 297–305.
Moorhouse RJ 1997. The diet of the North Island kaka
Hoare JM, O’Donnell CFJ, Wright EF 2010. Selection of
(
Nestor meridionalis septentrionalis) on Kapiti Island.
indicator species for State of the Environment reporting:
New Zealand Journal of Ecology 21: 141–152.
a case study from New Zealand. Pacific Conservation
Murphy EC, Clapperton BK, Bradfield PMF, Speed HJ
Biology 16: 76–82.
1998. Effects of rat-poisoning operations on abundance
Hutto RL, Young JS 2003. On the design of monitoring
and diet of mustelids in New Zealand podocarp forests.
programs and the use of population indices: a reply to
New Zealand Journal of Zoology 25: 315–328.
Ellingson and Lukacs. Wildlife Society Bulletin 31:
Murphy EC, Robbins L, Young JB, Dowding JE 1999.
903–910.
Secondary poisoning of stoats after an aerial 1080 operation
Innes J, Nugent G, Prime K, Spurr EB 2004. Responses of
in Pureora Forest, New Zealand. New Zealand Journal of
kukupa (
Hemiphaga novaeseelandiae) and other birds to
Ecology 23: 175–182.
mammal pest control at Motatau, Northland. New Zealand
Notman P 1989. A review of invertebrate poisoning by
Journal of Ecology 28: 73–81.
compound 1080. New Zealand Entomologist 12: 67–71.
Innes J, Kelly D, Overton JMcC, Gillies C 2010. Predation
Nugent G, Whitford J, Sweetapple P, Duncan R, Holland P
and other factors currently limiting New Zealand forest
2010. Effect of one-hit control on the density of possums
birds. New Zealand Journal of Ecology 34: 86–114.
(
Trichosurus vulpecula) and their impacts on native forests.
Johnson DH 2008. In defense of indices: the case of bird
Science for Conservation 304. Wellington, Department of
surveys. Journal of Wildlife Management 72: 857–868.
Conservation. 64 p.
Jones HP, Tershy BR, Zavaleta ES, Croll DA, Keitt BS,
O’Donnell CFJ 1995. Possums as conservation pests:
Finkelstein ME, Howald GR 2008. Severity of the effects
proceedings of a workshop on possums as conservation
of invasive rats on seabirds: a global review. Conservation
pests organised by the Possum and Bovine Tuberculosis
Biology 22: 16–26.
Control National Science Strategy Committee.
King CM 1983. The relationships between beech (
Nothofagus
Christchurch, New Zealand, 29-30 November 1994.
sp.) seedfall and populations of mice (
Mus musculus), and
Wellington, Department of Conservation. 81 p.
the demographic and dietary responses of stoats (
Mustela
O’Donnell CFJ 1996a. Predators and the decline of
erminea), in three New Zealand forests. Journal of Animal
New Zealand forest birds: an introduction to the hole-
Ecology 52: 141–166.
nesting bird and predator programme. New Zealand
King CM, O’Donnell CFJ, Phillipson SM 1994. Monitoring
Journal of Zoology 23: 213–219.
and control of mustelids on Conservation lands. Part
O’Donnell CFJ 1996b. Monitoring mohua (yellowhead)
2. Field and workshop guide. DOC Technical Series 4.
populations in the South Island, New Zealand, 1983-93.
Wellington, Department of Conservation. 36 p.
New Zealand Journal of Zoology 23: 221–228.
Larsen FW, Bladt J, Rahbek C 2007. Improving the performance
O’Donnell CFJ, Dilks PJ 1986. Forest birds in South Westland:
of indicator groups for the identification of important
status, distribution and habitat use. New Zealand Wildlife
areas for species conservation. Conservation Biology
Service Occasional Publication No. 10. 179 p.
21: 731–740.
O’Donnell CFJ, Dilks PJ 1989. Sap feeding by the kaka
Lawler JJ, Denis W, Sifneos JC, Master LL 2003. Rare species
(
Nestor meridionalis) in South Westland, New Zealand.
and the use of indicator groups for conservation planning.
Notornis 36: 65–71.
Conservation Biology 17: 875–882.
O’Donnell CFJ, Dilks PJ 1994. Foods and foraging of
Link WA, Cam E, Nichols JD, Cooch EG 2002. Of
BUGS and
forest birds in temperate rainforest, South Westland,
birds: Markov chain Monte Carlo for hierarchical modeling
New Zealand. New Zealand Journal of Ecology 18:
in wildlife research. Journal of Wildlife Management 66:
87–107.
277–291.
O’Donnell CFJ, Phillipson SM 1996. Predicting the incidence
Lloyd BD, McQueen SM 2002. Measuring mortality in short-
of mohua predation from the seedfall, mouse, and predator
tailed bats (
Mystacina tuberculata) as they return from
fluctuations in beech forests. New Zealand Journal of
foraging after an aerial 1080 possum control operation.
Zoology 23: 287–293.
New Zealand Journal of Ecology 26: 53–59.
O’Donnell CFJ, Dilks PJ, Elliott GP 1996. Control of a stoat
McLennan JA, Potter MA, Robertson HA, Wake GC, Colbourne
(
Mustela erminea) population irruption to enhance mohua
R, Dew L, Joyce L, McCann AJ, Miles J, Miller PJ, Reid
(yellowhead) (
Mohoua ochrocephala) breeding success
J 1996. Role of predation in the decline of kiwi,
Apteryx
in New Zealand. New Zealand Journal of Zoology 23:
spp, in New Zealand. New Zealand Journal of Ecology
279–286.
20: 27–35.
Payton I 2000. Damage to native forests. In: Montague TL ed.
Miller CJ, Anderson S 1992. Impacts of aerial 1080 poisoning on
The brushtail possum: biology, impact and management
the birds of Rangitoto Island, Hauraki Gulf, New Zealand.
of an introduced marsupial. Lincoln, Manaaki Whenua
New Zealand Journal of Ecology 16: 103–107.
Press. Pp. 111–125.
Miskelly CM, Dowding JE, Elliott GP, Hitchmough RA,
Powlesland RG, Wills DE, August ACL, August CK 2003.
Powlesland RG, Robertson HA, Sagar PM, Scofield RP,
Effects of a 1080 operation on kaka and kereru survival
140
New Zealand Journal of Ecology, Vol. 36, No. 2, 2012
and nesting success, Whirinaki Forest Park. New Zealand
Sweetapple PJ, Nugent G, Whitford J, Knightbridge PI 2002.
Journal of Ecology 27: 125–137
Mistletoe (
Tupeia antarctica) recovery and decline
R Development Core Team 2010. R: a language and
following possum control in a New Zealand forest.
environment for statistical computing. Vienna, Austria,
New Zealand Journal of Ecology 26: 61–71.
R Foundation for Statistical Computing. http://www.R
Thompson WL, White GC, Gowan C 1998. Monitoring
project.org.
vertebrate populations. San Diego, CA, Academic Press.
Rose AB, Pekelharing CJ, Platt KH, O’Donnell CFJ, Hall
365 p.
GMJ 1990. Impact of brush-tailed possums on forest
Tognelli MF 2005. Assessing the utility of indicator groups for
ecosystems, South Westland Forest Research Institute
the conservation of South American terrestrial mammals.
Contract Report FWE 90/52. Wellington, Department of
Biological Conservation 121: 409–417.
Conservation. 35 p.
Towns DR, Atkinson IAE, Daugherty CH 2006. Have
Royle JA, Dorazio RM 2006. Hierarchical models of animal
the harmful effects of introduced rats on islands been
abundance and occurrence. Journal of Agricultural,
exaggerated? Biological Invasions 8: 863–891.
Biological and Environmental Statistics 11: 249–263.
Urlich SC, Brady PJ 2005. Benefits of aerial 1080 possum
Sadleir R 2000. Evidence of possums as predators of native
control to tree fuchsia in the Tararua Range, Wellington.
animals. In: Montague TL ed. The brushtail possum:
New Zealand Journal of Ecology 29: 299–309.
biology, impact and management of an introduced
Veltman C 2000. Do native wildlife benefit from possum
marsupial. Lincoln, Manaaki Whenua Press. Pp. 126–131.
control? In: Montague TL ed. The brushtail possum:
Saunders A, Norton DA 2001. Ecological restoration at
biology, impact and management of an introduced
Mainland Islands in New Zealand. Biological Conservation
marsupial. Lincoln, Manaaki Whenua Press. Pp. 241–250.
99: 109–119.
Wardle JA 1984. The New Zealand beeches: ecology,
Spurr EB 1979. A theoretical assessment of the ability of bird
utilisation and management. Wellington, New Zealand
species to recover from an imposed reduction in numbers,
Forest Service. 447 p.
with particular reference to 1080 poisoning. New Zealand
Warman LD, Forsyth DM, Sinclair ARE, Freemark K, Moore
Journal of Ecology 2: 46–63.
HD, Barrett TW, Pressey RL, White D 2004. Species
Sweetapple PJ 2008. Spatial variation in impacts of brushtail
distributions, surrogacy, and important conservation
possums on two Loranthaceous mistletoe species.
regions in Canada. Ecology Letters 7: 374–379.
New Zealand Journal of Ecology 32: 177–185.
Westbrooke IM, Powlesland RG 2005. Comparison of impact
between carrot and cereal 1080 baits on tomtits (
Petroica
macrocephala). New Zealand Journal of Ecology 29:
Editorial Board member: Craig Barnett
143–147.
Received 18 August 2011; accepted 13 October 2011